This was originally made by Hirokazu Kobayashi but not copied from an online journal site to avoid copyright infringement.
The following Greek character used may be correctly displayed with Western fonts such as "Helvetica", "Times", etc.: "µ" for "micro".
Leaf-specifically expressed genes for polypeptides destined for chloroplasts with
domains of sigma70 factors of bacterial RNA polymerases in Arabidopsis thaliana
FOOTNOTES
Abbreviations: RNAP, RNA polymerase; SLF, sigma-like factor; Rubisco, ribulose-1,5
bisphosphate carboxylase/oxygenase; RT, reverse transcription;EST, expressed
sequence tag; GFP, green fluorescent protein.
Data deposition: The sequences reported in this paper have been deposited in the
GenBank database (accession nos. AB004820, AB004821, and AB004822 for SIG1, SIG2, and SIG3, respectively).
ABSTRACT
Genes for sigma-like factors of bacterial-type RNA polymerase have not been
characterized from any multicellular eukaryotes, although they probably play a crucial
role in the expression of plastid photosynthesis genes. We have cloned three distinct
cDNAs, designated SIG1, SIG2, and SIG3, for polypeptides possessing amino acid
sequences for domains conserved in sigma70 factors of bacterial RNA polymerases
from the higher plant Arabidopsis thaliana. Each gene is present as one copy per
haploid genome without any additional sequences hybridized in the genome.
Transient expression assays using green fluorescent protein demonstrated that N
terminal regions of the SIG2 and SIG3 ORFs could function as transit peptides for
import into chloroplasts. Transcripts for all three SIG genes were detected in leaves
but not in roots, and were induced in leaves of dark-adapted plants in rapid response
to light illumination. Together with results of our previous analysis of tissue-specific
regulation of transcription of plastid photosynthesis genes, these results indicate that
expressed levels of the genes may influence transcription by regulating RNA
polymerase activity in a green tissue-specific manner.
INTRODUCTION
The chloroplast is a semi-autonomous organelle whose genetic information is
encoded in the nuclear and plastid genomes. The plastid genome encodes genes for
photosynthesis, as well as genes for housekeeping functions such as protein
synthesis. There is evidence that photosynthesis genes are transcribed by a
multimeric Escherichia coli-type RNA polymerase (RNAP), and that housekeeping
genes are transcribed by a monomeric T7 or T3 bacteriophage-type RNAP (1). The -10
and -35 sequences, 5'-TATAAT-3' and 5'-TTGACA-3', respectively, in promoters of
many plastid genes (2) are recognized by the E. coli-type RNAP (3). The E. coli RNAP
is composed of a core complex of alpha, beta, and beta' subunits and one of a variety
of sigma factors, the principal one being sigma70, which is capable of binding to the
10 and -35 sequences (4-6). Determination of the complete nucleotide sequences of
plastid genomes from liverwort (7), tobacco (8), rice (9), and other plants has resulted
in finding of genes, rpoA, rpoB, and rpoC, probably encoding alpha, beta, and beta'
subunits, respectively, of a plastid RNAP. In higher plants, rpoC is duplicated, rpoC1
for beta' subunit and rpoC2 for beta" subunit. Amino acid sequences deduced from
maize plastid genes, rpoC2, rpoB, rpoC1, and rpoA, have proved to correspond to
those determined chemically of 180-, 120-, 78-, and 38-kDa polypeptides,
respectively, of highly purified maize plastid RNAP (10, 11).
No homolog for the bacterial-like sigma factor of RNAP has been detected in the
plastid genomes so far sequenced. However, some reports indicate that plastid sigma
like factors (SLFs) exist in higher plants. Antibodies against an E. coli sigma70
homolog from the cyanobacterium Anabaena sp. PCC7120 have cross-reacted with
polypeptides in purified plastid RNAPs frommaize and rice (12). In spinach 90- and 33
kDa polypeptides have been identified in a plastid RNAP that are immunologically
related to the sigma70 factor of E. coli RNAP, and these proteins bound to the
promoter of gene for the large subunit of ribulose-1,5-bisphosphate
carboxylase/oxygenase (Rubisco) (rbcL) and correctly initiated transcription in
association with RNAP core enzyme from spinach (13). Functionally distinct 67-, 52-,
and 29-kDa SLFs have been identified in mustard, and none of them alone bind to
DNA, but they confer enhanced binding to promoters and transcriptional activity on
RNAP core enzyme (14, 15). These experiments suggest that genes for homologs of
the E. coli sigma70 factor exist and are encoded in the nuclear genome. Nuclear
genes for such sigma factors have recently been reported in the red alga Cyanidium
caldarium (16, 17) but not in any multicellular eukaryotes.
We have demonstrated that expression of plastid photosynthesis genes is reduced
predominantly at the level of transcriptional initiation in nonphotosynthetic tissues of
Arabidopsis thaliana, indicating the importance of regulation through RNAP activity in
plastids (18). Therefore, we thought that cloning of genes for SLFs from multicellular
eukaryotes was needed for complete understanding of the regulatory mechanism. We
report here three cDNAs for putative sigma factors from the higher plant A. thaliana.
Amino acid sequences deduced from the cloned cDNAs have high homology to
conserved regions of the bacterial sigma70 factor family.
MATERIALS AND METHODS
Plant Materials.
A. thaliana ecotype Columbia was grown on vermiculite for 4 weeks at 22oC under 16
hr light / 8-hr dark. A. thaliana was also grown on a Murashige-Skoog (MS) agar
medium (19) without sugar at 22oC in continuous light at 3,000 lux for 3 weeks until
the growth stage of generation of 8 rosette leaves, and employed for reverse
transcription (RT)-PCR. Tobacco (Nicotiana tabacum cv. Petit Havana) was grown on a
MS agar medium (19) containing 3% sucrose at 28oC under continuous light at 4,000
lux.
Preparation of Nucleic Acids.
Total cellular DNA was prepared from leaves with cetyltriethylammonium bromide
(CTAB) (20), and RNA was isolated from leaves and roots with a Total RNA Separator
Kit (CLONTECH) according to the manufacturer's instructions. Poly(A)+ RNA fraction
was recovered from the total RNA by using Oligotex-dT30 (Takara, Otsu, Japan). Total
cellular RNA was also prepared by Isogen (NipponGene, Toyama, Japan) and treated
with RQ1 (RNase-free DNase, Promega) following the suppliers' instructions for
employment for RT-PCR.
Screening of cDNAs for sigma70 Homologs.
The amino acid sequence GYKFSTYAMWWIRQAITRSIAD, which is responsible for
DNA melting and recognition of the -10 sequence in bacterial promoters, was highly
conserved in sigma70 factors in bacteria. The sequence was subjected to homology
search in the database of A. thaliana expressed sequence tags (ESTs). Three EST
clones were found with accession numbers stock numbers at the Arabidopsis
Biological Resource Center (ABRC), number of nucleotides reported, including
unidentified ones, N65838 [240C23T7, 538 bp], T88387 [155H23T7, 389 bp], and
N97044 [242P3T7, 535 bp]. DNA fragments corresponding to the first two of these
clones were amplified by PCR from total cellular DNA from A. thaliana with primers
designed on thebasis of their nucleotide sequences in the EST database. The PCR
products were inserted into pT7Blue-T (Novagen) and sequenced to confirm the
inserts. These two inserts, as well as a SalI-XhoI fragment from 242P3T7A, were
labeled with alpha-[32P]dCTP by using a Random Primer DNA Labeling Kit (Takara),
and used to screen a lambda ZAP cDNA library (4 X 105 plaque-forming units) of A.
thaliana leaves, which was constructed by using the ZAP-cDNA Synthesis Kit
(Stratagene). The inserts of positively hybridizing clones were sequenced by an
Applied Biosystems 373A DNA sequencer.
Hybridization of Nucleic Acids.
Total cellular DNA digested with PstI or EcoRI was electrophoresed in 1% agarose
gels, and poly(A)+ RNA was electrophoresed in 1.2% agarose gels containing 0.66 M
formaldehyde (21). The nucleic acids were transferred to nylon membranes (Hybond
N+, Amersham) with 0.4 M NaOH for DNA and 20 X SSC for RNA (1 X SSC = 0.15 M
NaCl / 0.015 M sodium citrate, pH 7.0). The prehybridization and hybridization were
performed at 65oC (21). Gene-specific DNA probes were labeled with alpha [32P]dCTP by using a Random Primer DNA Labeling Kit (Takara). Membranes were
washed with 2 X SSC containing 0.1% SDS for 30 min at 65oC, and with 0.5 X SSC
plus 0.1% SDS under the same conditions. Radioactivity on the membranes was
detected by BAS2000 Bio-Imaging Analyzer (Fujix, Tokyo).
RT-PCR.
Total cellular RNA prepared from A. thaliana was treated with RQ1 DNase, following
the supplier's instructions. RT-PCR was performed with approx. 1 µg of the total RNA
and oligo(dT)12-18 in 21 µl of reaction mixture by using the SuperScript
Preamplification System for First Strand cDNA Synthesis (GIBCO/BRL). An aliquot (2
µl for SIG genes, described in Results, or 1 µl for ACT2 encoding actin 2) from the
reaction mixture was further subjected to PCR with AmpliTaq (Perkin-Elmer). Primers
for PCR of SIG1, SIG2, and SIG3 (see Results) were designed to amplify 395-, 254-,
and 342-bp DNA fragments, respectively, on the basis of nucleotide sequences of
cDNAs determined in this investigation: 5'-CCTCCGAGGTCGCTAGACCAG-3' and 5'
GCTCAGGTGGGTGGGTTCTATGC-3' for SIG1, 5'-GGTCTCCCTGGAGAAACTCATC
3' and 5'-CTCCAGCGCCACAAGCCCTACC-3' for SIG2, and 5'
GATTCGAGCAGCTAACCAATGCC-3' and 5'-GAGGTCTTCGGCGTTGGGTTGG-3' for
SIG3. ACT2 (22) was also examined as an internal standard with primers, 5'
GAAGATTAAGGTCGTTGCACCACCTG-3' and 5 '
ATTAACATTGCAAAGAGTTTCAAGGT-3', to amplify the 477-bp DNA fragment with
cDNA or the 563-bp one with genomic DNA, if any, contaminating the RNA fractions.
Transient Expression of Chimeric Genes.
DNA fragments encoding the first 83 and 89 amino acid residues of the ORFs of SIG2
and SIG3 cDNAs, respectively, were amplified by PCR with pairs of synthetic
oligonucleotides containing new NcoI sites (italicized): 5'
TTCCATGGCTACTGCAGCTG-3' and 5'-TTCCATGGTAGAAGCAACATCATC-3' for
SIG2, and 5'-TTCCATGGCTTCCTTCA-3' and 5'-TTCCATGGATAGAAACGACC-3' for
SIG3. Two PCR products were digested with NcoI and inserted into the NcoI site of
CaMV35S-sGFP(S65T)-nos [pUC18, harboring a synthetic gene for improved green
fluorescent protein sGFP(S65T), hereafter referred to as "GFP"] driven by cauliflower
mosaic virus 35S promoter and NOS terminator (23), resulting in two chimeric GFP
constructs, SIG2-GFP and SIG3-GFP. The synthetic GFP gave approx. 100 times
higher fluorescent signal upon excitation with 488-nm light in comparison with that of
the native GFP (23).The chimeric constructs were introduced into tobacco leaf cells by
using Biolistic PDS-1000/He (Bio-Rad). Gun parameters employed were as follows:
pressure rupture disks rated at 1,100 psi (7.58 MPa), a vacuum at 27 inches of Hg
(reaching 9.83 kPa), distance to target tissues 6 cm, and gold particles 1.0 µm in
diameter. After bombardment, cells were incubated for 20 hr at 28oC under continuous
light, and observed by using the Bio-Rad MRC-1024 Confocal Imaging System (480
X).
RESULTS
Three Distinct cDNAs for Polypeptides with Conserved Domains in sigma
Factors.
Three cDNA clones (reported partial sequences of 538 bp, 389 bp, and 535 bp,
including unidentified nucleotides) in the EST database of A. thaliana contain animo
acid sequences that are highly homologous to regions for melting DNA and
recognizing the -10 promoter sequence of bacterial sigma factors. The sequences in
these clones were employed as probes to screen a leaf cDNA library from A. thaliana,
and positively hybridizing clones, 6, 9, or 7, respectively, for these three ESTs, were
obtained from the library of 4 X 105 plaque-forming units. Three cDNAs,
corresponding to stock numbers 240C23T7, 155H23T7, and 242P3T7, were
designated SIG1, SIG2, and SIG3, respectively. The largest ORFs of SIG1, SIG2, and
SIG3 coded for polypeptides with 572, 502, and 571 amino acids, respectively.
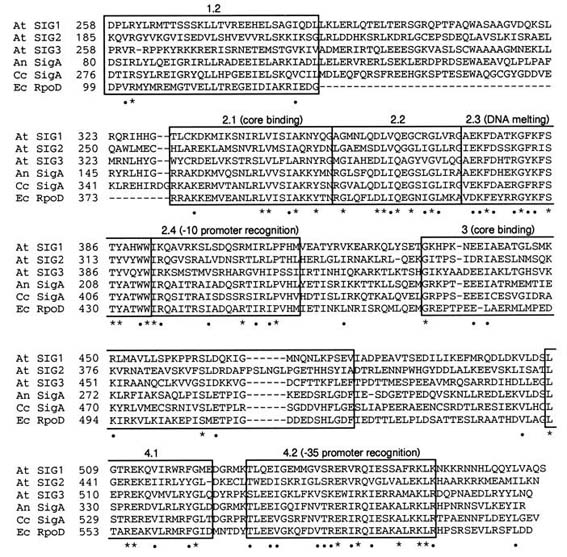
Similarity of Functional Domains in sigma Factors.
Amino acid sequences deduced from SIG1, SIG2, and SIG3 were compared with
sigma factors from E. coli (24), Anabaena sp. PCC7120 (25), and Cyanidium
caldarium (17) (Fig. 1). The comparison showed that all ORFs of SIG genesare highly
conserved in two major regions (designated regions 2 and 4) in bacterial sigma factors
(26). Subregions 2.1, 2.3, 2.4, and 4.2 are involved in interaction with core enzyme,
DNA melting, recognition of -10 promoter sequence, and recognition of -35 promoter
sequence, respectively. The subregion 4.1 can be modeled as an amphipathic alpha
helix. Region 3 may contribute to binding to core RNAP. The functions of subregions
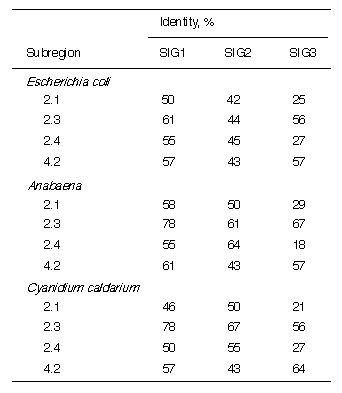
1.2 and 2.2 are unclear. Table 1 shows comparisons of SIG1, SIG2, and SIG3 with the
highly conserved subregions 2.1, 2.3, 2.4, and 4.2 from E. coli, Anabaena, and C.
caldarium. Among three SIGs, SIG1 has the highest similarity to bacterial sigma
factors, and SIG3 has the least similarity. Among the subregions of the SIGs, 2.3 is the
most conserved, while 2.1 and 2.4 are the least conserved, especially in SIG3 (18-29%). The subregion 4.2 in SIG2 is more diverse from bacterial sigma factors, showing
43% in spite of 57-61% in SIG1 and 57-64% in SIG3.
----------------------------------------------------

Fig. 1. Alignment of conserved domains in the deduced amino acid sequences of A.
thaliana SIG1 (At SIG1), SIG2 (At SIG2), and SIG3 (At SIG3) with those of SigA from
Anabaena sp. PCC 7120 (An SigA), SigA from C. caldarium (Cc SigA), and sigma70
factor encoded in rpoD from E. coli (Ec RpoD). Regions 1-4 in bacterial sigma factors
(5, 26) are boxed. Residues marked with an asterisk (*) or a dot (.) are identical or
similar, respectively, in all six sequences. The numbers between the product
designations and the amino acid sequences are those of amino acid residues. There
are 246 amino acids in a gap between subregion 1.2 and subregion 2.1 in E. coli
RpoD.
----------------------------------------------------
----------------------------------------------------
Table 1. Identity of A. thaliana SIG1, SIG2, and SIG3 to other factors in the sigma70
family in major conservative regions
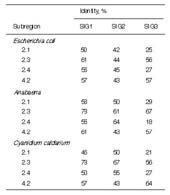
See Fig. 1 for sequences of the sigma factors.
----------------------------------------------------
Numbers and Expression of the SIG Genes.
Total cellular DNA from A. thaliana was digested with PstI or EcoRI and subjected to
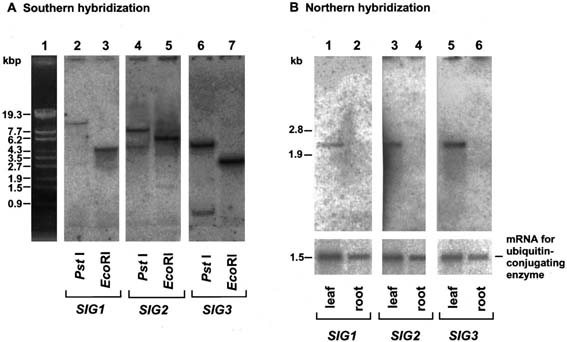
Southern hybridization using SIG1, SIG2, and SIG3 as probes (Fig. 2A). One band
was generated in each case except for SIG2 and SIG3 digested with PstI (lanes 4 and
6). Multiple bands in these lanes resulted from the presence of PstI sites in these two
SIG genes. Therefore, SIG1, SIG2, and SIG3 are probably single-copy genes in A.
thaliana.
We have already demonstrated that suppressed expression of the plastid genome
encoded photosynthesis genes in roots of A. thaliana is due predominantly to
transcriptional regulation (18). To examine the possibility that there is an involvement
of SIGs in the transcriptional regulation, levels of transcripts for the SIG genes in RNAs
from leaves and roots were investigated by Northern hybridization. All transcripts for
SIG1, SIG2, and SIG3 were detected in leaves, but barely in roots (Fig. 2B). This
finding suggests that expressed levels of SIG genes correlate with transcriptional
activities of plastid photosynthesis genes evaluated by run-on transcription (18).
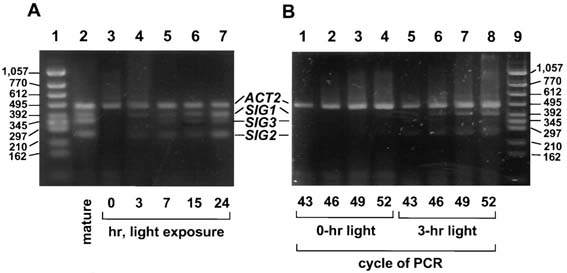
Light induction of SIG transcripts in leaves of dark-adapted plants was examined by
RT-PCR. Transcripts for ACT2 (encoding actin 2), which is reported to be constitutively
expressed (22), were determined as internal standards both for proving no
contamination with genomic DNA in the RNA fractions and estimating the amounts of
SIG transcripts. Expression of all three SIG genes was obviously under light control
(Fig. 3). SIG1 and SIG2 transcripts were induced during 3-hr illumination (Fig. 3B) and
seemed to reach a plateau between 7 and 15 hr in the light induction (Fig. 3A).
----------------------------------------------------
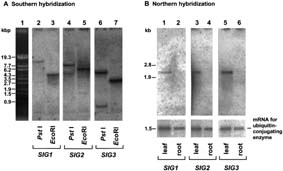
Fig. 2. Southern and Northern blot analyses of SIG genes. (A) Southern hybridization
with SIG gene probes of total cellular DNA (8 µg perlane) digested with PstI (lanes 2,
4, and 6) and EcoRI (lanes 3, 5, and 7). DNA in lanes 2 and 3, lanes 4 and 5, and
lanes 6 and 7, was hybridized with 1.7-kb EcoRI fragment from SIG1 cDNA, 1.6-kb
XbaI fragment from SIG2 cDNA, or 1.2-kb XhoI fragment from SIG3 cDNA,
respectively. (B) Northern hybridization with SIG probes of total poly(A)+ RNA (0.5-1.0
µg per lane) from leaves (lanes 1, 3, and 5) and roots (lanes 2, 4, and 6). The cDNA for
ubiquitin-conjugating enzyme, which is constitutively expressed (M.S., Y.N., and H.K.,
unpublished results), was used as a probe as an internal control for equivalence of
amounts of RNA loaded into lanes. Each of the SIG genes gives rise to approx. 2.3-kb
mRNA.
----------------------------------------------------
----------------------------------------------------
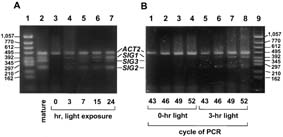
Fig. 3. Time-sequential changes of transcripts for SIG genes in dark-adapted plants
after exposure to light. (A) A. thaliana grown for 3 weeks was incubated in the dark for
3 days or maintained under the same light conditions for plants indicated as "mature."
Leaves of individual dark-adapted plants were harvested at 0, 3, 7, 15, and 24 hr after
initiation of the exposure to light at 3,000 lux. RT-PCR products were subjected to
electrophoresis after PCR cycles when intensities of signals for ACT2 were
unsaturated and comparable among RNA fractions harvested at different illumination
times. RT-PCR products (5 µl from 50 µl of reaction mixture) were electrophoresed in
3% agarose gel (Agarose H, NipponGene), stained with SYBR green I (FMC
BioProducts), and observed by FluorImager SI (Molecular Dynamics). (B) RT-PCR
products of 0- and 3-hr illumination after indicated PCR cycles were electrophoresed
and stained with SYBR green I.
----------------------------------------------------
Destination of SIG Gene Products for Chloroplasts.
Transit peptides can be predicted by PSORT, a program for protein sorting
(http://psort.nibb.ac.jp/index.html) (27). The N-terminal regions of SIG2 and SIG3 (Fig.
4) were predicted to be transit peptides at probability higher than 90%. These N
terminal sequences are enriched with Ser and Thr, a property of transit peptides (28).
However, the N-terminal region of SIG1 ORF (Fig. 4) was less likely to be a transit
peptide.
We have tested whether N-terminal regions of SIGs can function as transit peptides for
plastid-targeting by transient expression assays with animproved GFP. We made two
constructs in which peptides composed of the first 83 amino acids of SIG2 ORF and of
the first 89 residues of SIG3 ORF (Fig. 4) were fused to the N terminus of GFP (SIG2
GFP and SIG3-GFP) and placed under the control of cauliflower mosaic virus 35S
promoter. Tobacco leaves were bombarded with one of these constructs. The GFP
alone (23) and a chimeric construct of GFP with the transit peptide of small subunit of
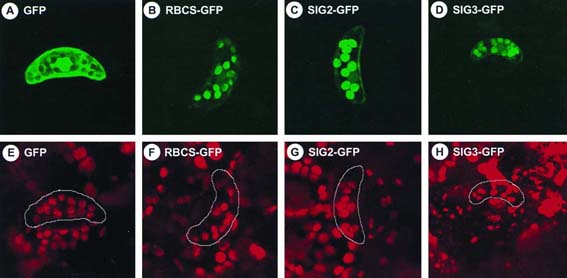
Rubisco [RBCS-GFP, as a positive control (23)] were used. Fig. 5 shows results with
tobacco guard cells, in which it is easier to visualize GFP accumulated in chloroplasts.
Green fluorescence was observed only in chloroplasts of the guard cells (Fig. 5 B, C,
and D), where chloroplasts were identified by red fluorescence of chlorophyll (Fig. 5 F,
G, and H). Without these N-terminal sequences, GFP alone was not localized in
chloroplasts (Fig. 5 A and E). We obtained similar results with leaves of A. thaliana
(data not shown). These data suggest that the N-terminal regions of SIG2 and SIG3
ORFs can function as transit peptides for import into chloroplasts. We could not confirm
the function of N-terminal region of SIG1 as a transit peptide by transient expression
with GFP (data not shown).
----------------------------------------------------

Fig. 4. N-terminal sequences of SIG1, SIG2, and SIG3. The first 90 amino acids in the
ORFs of SIG1 (SIG1), SIG2 (SIG2), and SIG3 (SIG3) cDNAs are represented. Ser and
Thr residues are indicated in boldface.
----------------------------------------------------
----------------------------------------------------
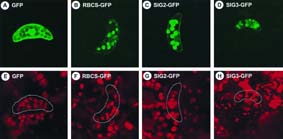
Fig. 5. Localization of GFPs fused to N-terminal regions of SIG2 and SIG3. GFP fusion
constructs with the N-terminal regions of SIG2 ORF (SIG2-GFP, C and G) and SIG3
ORF (SIG3-GFP, D and H), and the transit peptide of the small subunit of Rubisco (23)
(RBCS-GFP, B and F), as well as GFP alone (23) (GFP, A and E), were introduced into
tobacco leaves by particle bombardment. Guard cells were observed by using the
MRC-1024 Confocal Imaging System (480 X) with excitation at 488 nm and emission
at 520 nm (A-D), as well as excitation at 647 nm and emission at 666 nm (E-H). The
same objects are shown in each pair of upper and lower panels.
----------------------------------------------------
DISCUSSION
When the complete nucleotide sequence of tobacco plastid genome was determined
in 1986 (8), genes for all subunits of E. coli-type RNAP except for SLFs were found in
the plastid genome. In spite of thorough effort to identify genes for SLFs in plants, they
were not cloned until 1996, when the genes were isolated from the unicellular red alga
Cyanidium caldarium by two research groups (16, 17). In the investigation reported
here, putative genes for sigma factors from multicellular eukaryotes have now been
characterized. It took over a decade to obtain clones for SLFs from higher plants for
the following reasons: (i) low contents of mRNA, one in 40,000-70,000 molecules of
total poly(A)+ RNA in leaves of A. thaliana as estimated from the frequency of cloning
in this investigation; (ii) homology conserved but at degree nonhybridizable to
bacterial ones; and (iii) possible interference by the gene products with the growth of
the E. coli used for cloning (data not shown).
The ORFs of SIG1, SIG2, and SIG3 encode polypeptides consisting of 572, 502, and
571 amino acids, respectively, with calculated molecular masses of 64, 56, and 65
kDa. Although these are precursors with transit peptides, they are close to 67 kDa
(SLF67) and 52 kDa (SLF52) of mustard SLFs associated with the activity of sigma
factors (16, 29) and to 64-kDa peptides from maize and rice that immunochemically
cross-reacted with antibodies against a sigma factor from Anabaena (12). SIG1 among
the three SIGs most resembles bacterial sigma factors and has more than 50% identity
to four major conservative subregions---2.1, 2.3, 2.4, and 4.2 ---in each of the sigma
factors from E. coli, Anabaena, and C. caldarium except for the subregion 2.1 in C.
caldarium. SIG2 has lower identity in its subregion 4.2 for -35 promoter recognition,
and SIG3 has significantly low identity in both its subregions 2.1 and 2.4 for DNA
melting and -10 promoter recognition, respectively. The differences inamino acid
sequences among the subregions of the three SIGs may contribute to fine regulation
of transcription of distinct target genes.
We have already demonstrated that expression of plastid photosynthesis genes, rbcL
for large subunit of Rubisco, psbA for D1 protein in photosystem II reaction center, and
atpB/E for beta and epsilon subunits of coupling factor 1, is remarkably suppressed by
transcriptional regulation in roots in A. thaliana (18). Northern analysis of SIG1, SIG2,
and SIG3 gene expression showed that their transcripts were few in roots but were
present in leaves (Fig. 2B). This observation suggests that changes of SIG levels could
regulate the expression of plastid photosynthesis genes in a tissue-specific manner.
The light induction of transcripts for SIG genes (Fig. 3) probably precedes induction of
transcripts of plastid photosynthesis genes such as rbcL (data not shown). Therefore, it
is very probable that generation of all SIG gene products results in the initiation of
transcription of photosynthesis genes after their transport into plastids and association
with RNAP core enzyme in plastids. We think that the present investigation has
opened a way to study not only the biological significance of each of the three distinct
SIGs but also the regulation of their tissue-specific expression.
REFERENCES
- Allison, L. A., Simon, L. D. & Maliga, P. (1996) EMBO J. 15, 2802-2809.
- Kung, S. D. & Lin, C. M. (1985) Nucleic Acids Res. 13, 7543-7549.
- Igloi, G. L. & Kel, H. (1992) Crit. Rev. Plant Sci. 10, 525-558.
- Burgess, R., Travers, A., Dunn, J. & Bautz, E. (1969) Nature (London) 221, 43-46.
- Helmann, J. D. & Chamberlin, M. J. (1988) Annu. Rev. Biochem. 57, 839-872.
- Busby, S. & Ebright, R. H. (1992) Cell 79, 743-746.
- Ohyama, K., Fukuzawa, H., Kohchi, T., Shirai, H., Sano, T., Sano, S., Umesono, K.,
Shiki, Y., Takeuchi, M., Chang, Z., Aota, S., Inokuchi, H. & Ozeki, H. (1986) Nature
(London) 322, 572-574.
- Shinozaki, K., Ohme, M., Tanaka, M., Wakasugi, T., Hayashida, N., Matsubayashi,
T., Zaita, N., Chunwongse, J., Obokata, J., Yamaguchi-Shinozaki, K., Ohto, C.,
Torazawa, K., Meng, B., Sugita, M., Deno, H., Kamogashira, T., Yamada, K., Kusuda,
J., Takaiwa, F., Kato, A., Tohdoh, N., Shimada, H. & Sugiura, M. (1986) EMBO J. 5,
2043-2049.
- Hiratsuka, J., Shimada, H., Whittier, R., Ishibashi, T., Sakamoto, M., Mori, M.,
Kondo, C., Honji, Y., Sun, C.-R., Meng, B.-Y., Li, Y.-Q., Kanno, A., Nishizawa, Y., Hirai,
A., Shinozaki, K. & Sugiura, M. (1989) Mol. Gen. Genet. 217, 185-194.
- Hu, J. & Bogorad, L. (1990) Proc. Natl. Acad. Sci. USA 87, 1531-1535.
- Hu, J., Troxler, R. F. & Bogorad, L. (1991) Nucleic Acids Res. 19, 3431-3434.
- Troxler, R. F., Zhang, F., Hu, J. & Bogorad, L. (1994) Plant Physiol. 104, 753-759.
- Lerbs, S., Bräutigam, E. & Mache, R. (1988) Mol. Gen. Genet. 211, 459-464.
- Bülow, S. & Link, G. (1988) Plant Mol. Biol. 10, 349-357.
- Tiller, K., Eisermann, A. & Link, G. (1991) Eur. J. Biochem. 198, 93-99.
- Liu, B. & Troxler, R. F. (1996) Proc. Natl. Acad. Sci. USA 93, 3313-3318.
- Tanaka, K., Oikawa, K., Kuroiwa, H., Kuroiwa, T. & Takahashi, H. (1996) Science
272, 1932-1933.
- Isono, K., Niwa, Y., Satoh, K. & Kobayashi, H. (1997) Plant Physiol. 114, 623-630.
- Murashige, T. & Skoog, F. (1962) Physiol. Plant. 15, 473-497.
- Rogers, S. O. & Bendich, A. J. (1985) Plant Mol. Biol. 5, 69-76.
- Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989) Molecular Cloning: A Laboratory
Manual (Cold Spring Harbor Lab. Press, Plainview, NY), 2nd Ed.
- An, Y. Q., Mcdowell, J. M., Huang, S. R., Mckinney, E. C., Chambliss, S. &
Meagher, R. B. (1996) Plant J. 10, 107-121.
- Chiu, W. L., Niwa, Y., Zeng, W., Hirano, T., Kobayashi, H. & Sheen, J. (1996) Curr.
Biol. 6, 325-330.
- Burton, Z., Burgess, R. R., Lin, J., Moore, D., Holder, S. & Gross, C. A. (1981)
Nucleic Acids Res. 9, 2889-2903.
- Brahamsha, B. & Haselkorn, R. (1991) J. Bacteriol. 173, 2442-2450.
- Lonetto, M., Gribskov, M. & Gross, C. (1992) J. Bacteriol. 174, 3843-3849.
- Nakai, K. & Kanehisa, M. (1992) Genomics 14, 897-911.
- Keegstra, K. & Olsen, L. J. (1989) Annu. Rev. Plant Physiol. Plant Mol. Biol. 40,
471-501.
- Tiller, K. & Link, G. (1993) Plant Mol. Biol. 21, 503-513.