
���ƗY�̘b
���� ���� �� (Prof. Tohru Kobayashi)���̔�����`�w������
���ԉ��w�������́A2009�N10����萶�B�����w����ՂƂ����������Ƀ��j���[�A�������B����́A�`�Ԋw�A���q�����w����g���Č̘̂A�����i��̈ێ��j���\�Ƃ����鐶�B���ۂ̊�{�������𖾂��A��̑��l���A�ۑS�ɍv�����邱�Ƃ�ڎw���B���ʂ͓��ɁA�u���Y���v�̖��Ɏ��g�݂����ƍl���Ă���B
���B�B�́A���Y�̊Ԃōł������ȍ��ق������g�D�ł���i�����Ɨ����j�A���B�����ɕs���ł��邱�Ƃ͌����܂ł��Ȃ��B��q����悤�ɁA���B�B�ł͐��B�B�����ߒ��ɂ����鐫�z�������̎Y�����s�Ȃ��A���炾�S�̂ɐ�����U������B�܂�A�����Ɨ����̕����i�������j�͌̂̎��Y������������ߒ��ōł��d�v�Ȍ��ۂł���B������A���B�B�̔����E�����A���z�������̍�p�A���s�����́u���Ɋ֘A���錻�ہv�͂�����������[�������N����B���Ɋւ�錻�ۂ́A��w�A��w�A�_�w�A���Y�w�A���Ȋw�ȂǗl�X�Ȍ�������Ɩ��ڂɊւ��Ȃ���A����̐i�W�����҂ł��錤������̈�ł���B
�͂��߂�
���i���Y���j�́A��ʂɌ̂Ƃ��Ă̐����Ӗ�����B�������q�g���܂ޑ��זE�����̌̂́A�̎��g�ł͉i���̐���������ł͂Ȃ��A���肪����B���ꂼ��̎���ێ�������ɂ́A���炩�̌`�Ō̂�A��������K�v�����邱�Ƃ͎����ł���B��ʓI�ɂ́A���זE�����̌̂Ƃ��Ă̘A�����́A�L�����B�i���Y���j�ɂ���ĕۏ���Ă���A����ɂ���̈ێ��A�ɉh���\�Ƃ�����B�̘̂A�����́A�B�ꎟ����ɂȂ���זE�ł��鐶�B�זE����������ߒ����o�ĕ��������z��q�ł��鐸�q�A���ɂ���ď��߂ĉ\�ƂȂ�B�����Đ��q�Ɨ����ɂ���ĂP�̎��ƂȂ�זE������J��Ԃ�����A�̂ւƕ������A������ւƂȂ���i�}�P�j�B
�̂̐��͑�ʂ����1)�A2)���B�B�̐������A3)���z�������ɂ�鐫���̗U���A�̎O�̉ߒ����琬�藧���Ă���B�P�j�F���̎��_�Ő����F�̂̑g�ݍ��킹�����܂�A�M���ނ̏ꍇXY�ŗY�AXX�Ŏ��ƂȂ�B�Q�j���B�B�i�����Ɨ����j�̐������F���B�B�͒���t�̈ꕔ���甭�����A���̗̈�͐����F�̂̑g�ݍ��킹�ɂ��������āA�Y�ł͐������A���ł͗�������������B���̌�A���������������Ɨ�������͐��z�������i���X�e���C�h�z�������j�Ƃ��Ă��ꂼ��A�j���z�������i�A���h���W�F���j�Ə����z�������i�G�X�g���W�F���j�������������B�R�j���z�������ɂ�鐫���̗U���F�����Ɨ����ŎY�����ꂽ���z�������͌��ǂ�ʂ��ėl�X�ȑg�D�ւƉ^��A���z��������e�́i�j���z��������e�̂Ə����z��������e�́j�Ɍ������A��p����B���z��������e�͓̂]�ʈ��q�Ƃ��āA���z�������̌����ɂ���ĕW�I��`�q�̓]�ʂ߂���B�܂�A�t�����B�튯��]�̐����̍\�z�ߒ��ɂ��A���z��������e�̂ɂ���Ē��߂�����`�q�������K�v�ƂȂ�B
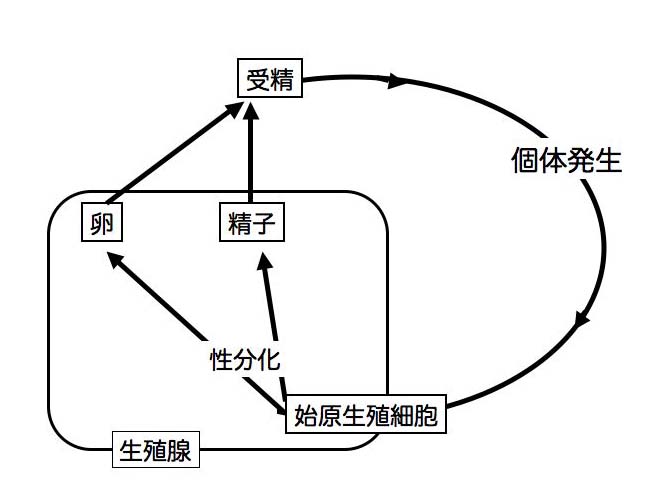
�̘̂A��������̈ێ��������炷�D���̌��ۂ��u���B�v�ł���D�Ғœ����́u���B�v�͎��Y���������L�����B�ł���A���̉ߒ��Ő�����A���������N����D
������Ɛ�����
���̌���l���͓�����ɂ���ĈقȂ�A���̗v������`��������푽�l�ł���B�܂��A���]�����铮���⎓�Y���̌��ۂ���������������������݂���B���������I��^���������B�B�i�����E�����j�Ƃ����튯�͏�ɑ��݂��A�Ғœ����ł͂��̐��B�B�̐��I�^���̂̐��̕\���^�̊�ՂƂȂ�B���Ȃ킿���̌���l���͑��l�ł��邪�A���̐��B�B�`���ߒ��ł͑��זE�������ʂ̕��q�@�\�������Ă��邱�Ƃ��\�z�����B�����A�ŋ߂̌������ʂ��琫����͎���ٓI�ȃX�C�b�`�ōs�Ȃ��Ă��邱�Ƃ����炩�ƂȂ��Ă���B�Ғœ����̐������`�q�́A1990�N�Ƀq�g�̐������`�q��SRY�Ɠ��肳�ꂽ�� (Sinclar et al., 1990)�A�M���ވȊO�̐Ғœ����ɂ�����SRY�̑�����`�q�̑��݂��m���߂錤�������͓I�ɍs�Ȃ��Ă������A����܂łɂ��̂悤�Ȉ�`�q�͌������Ă��Ȃ��B�ŋ߂̃��_�J�ōs�Ȃ�ꂽ�Ғœ����łQ�Ԗڂ̐������`�q�Fdmy�̓���̌����́A�������`�q�͓�������ٓI�ł��邱�Ƃm�Ɏ������idmy����Ɋւ���b�͑����Q�Ƃ��ꂽ��[1]�j�B
���B�B�̐������́A���Ȃ��Ƃ��Q�̈Ӗ����܂�ł���i�}2�j�B�P�́A���B�B�ɑ��݂��鐶�B�זE���A���q�ɂȂ�̂��A���ɂȂ�̂��i���B�זE�̐������j�B�����P�́A���B�B�̍\�����Y�^�ɂȂ�̂��i�����j�A���^�ɂȂ�̂��i�����j�Ƃ������Ƃł���i���B�B�̐������j�B���̂Q�̐������݂͌��ɋ������ċN�����Ă��邱�Ƃ��m���Ă���B���q�◑�ɕ�������匳�̐��B�זE�i�n�����B�זE�j�́A�����F�̂����Ғœ����ł́A���̍זE�i�̍זE�j�Ɠ��l��XX��������XY�̐��F�̍\���������A���Ȃ킿�A���̈�`�q�^�Ƃ��Ă̐��͑��݂��Ă���͂��ł���B�������A�����̏ꍇ�Ŏn�����B�זE�͂��̈�`�q�^�ɂ�炸�A��������̗��\���������Ă���B�n�����B�זE�����I���\���������Ƃ͉����Ғœ����ł͖��炩�ł���B���ނł͑����̎�Ŏ��R��ԁA���邢�͎����I�������Ƃ������������̕ω��Ő��]�����N���邱�Ƃ��m���Ă���B�܂��A���R��ԂŐ��]�����N��������A�Љ�I�Ȓn�ʂɂ���Đ��]�������肷����̂ł́A��`�I�Ȑ��Ƃ������̂������B�܂�A�������`�q�������ƍl�����A�����̎n�����B�זE�͐��I���\���ł��邱�ƂɂȂ�B���̂��Ƃ́A�����Ғœ����Ɍ��������Ƃł͂Ȃ��A�M���ނɂ����Ă������I�ɂ͐��I�Y���������Ă��邱�Ƃ��m���Ă���B�܂��A���̐��I�Y���͎n�����B�זE�Ɍ���ꂽ���Ƃł͂Ȃ��A��i����j�ł��鐶�B�B���\�����Ă���̍זE�ł����S���͂Ƃ������A���̐����������Ă��邱�Ƃ��Ӗ�����B
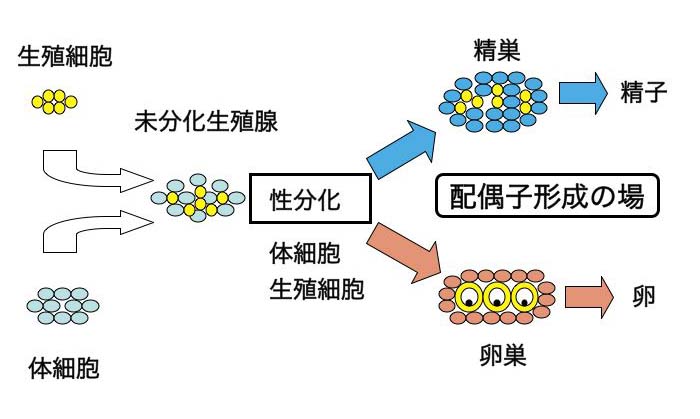
����������J��Ԃ��A�̂Ƃ��Ắu���炾����v�����Ă����ߒ��Ő��B�זE�i�n�����B�זE�j���`������A���B�B���`�������ꏊ�܂ňړ����Ă���D�����ŁA���I�ɖ������Ȗ��������B�B���`������.���������B�B�͈�`�q�^�ɏ]���āA���B�זE�A�̍זE������������D�������ɂ�萶�B�B�͐����A�����ւƕ����E���B����D
���I�Y��
���ނł́A�l�דI�ɐ��z�������i�����z��������j���z�������j�𓊗^���邱�Ƃɂ���ėe�Ղɖ{���̐��i��`�q�^�̐��FXX�AXY�j�Ƃ͋t�̐��ɐ��]�������邱�Ƃ��ł���B���ނ̂悤�ɐ��z���������^����ݐ��G�X�g���Q���̒��߂ɂ���ėe�Ղɐ��]�����N������̂ł́A���v�������B�B�̍זE�̐��I�Y���ɋ����֗^���邱�Ƃ���������Ă���[2]�B�܂��A��q�����悤�ɁA���ނł͎Љ�������̕ω��ɂ��A���n�̂ł����Ă����]������킪�����m���Ă���B�����ł́A����ȗ�ł͂��邪�A�M�ш�̃T���S�ʂɐ������鋛�ł݂��闼�����i�Y���玓�B������Y�j�̐��]���ɂ��Đ������悤�B��ʓI�ɒm���Ă���u������Y�v�i������n�j�A�u�Y���玓�v(�Y����n)�A���]���ɂ��Ă͑����Q�Ƃ��ꂽ��[1,2]�B
![�}3 ���Y�������ɐ��]������I�L�i���x�j�n�[](./images_24/fig_24-3_full.jpg)
- �I�L�i���x�j�n�[ (Trimma okinawae)
- ���̐��B�B�i���j�ł́A�����iO�j���傫���A�����̑�^�̗���זE���݂���.�Y�̐��B�B�i���j�ł͐����iT�j���傫���A���q���݂���D
- �召�̂Q���̎����y�A�Ŏ��炷��ƁA�T����ɂ͑傫�����̎����Y�ɐ��]������D�召�Q���̗Y���y�A�Ŏ��炷��ƁA�P�O����ɂ͏��������̗Y�����ɐ��]������D
�������̐��]��
�I�L�i���x�j�n�[�iTrimma okinawae�j�͑傫�Ȏ��Ə����Ȏ��������ɓ����Ɩ�T���ő傫�Ȏ������]�����A�Y�ƂȂ�B���̗Y�́A���������ɂ���ɑ傫�ȕʂ̗Y������ƍĂю��ɖ߂�B�����悤�ȏ������J��Ԃ��Ɖ��x�ł����]�����N����B����𗼕����̐��]���ƌĂ�[1]�B���̐��]���͎��o��L���[�ɂȂ��Ă���B�I�L�i���x�j�n�[�̐��]���ߒ��ɂ����Đ��s���̕ω��́A���]���̕����ɊW�Ȃ��A�R�O���ȓ��ɋN����B���B�B�̋@�\�I���]���́A�u������Y�ցv�͂T���A�u�Y���玓�ցv�͂P�O���Ŋ�������B���̋��͉�U�w�I�ɂ͎��Y�Ƃ��ɂP�̐����y�ї����i�v�Q�̐��B�B�j�����B���ꂪ�A�������̐��]�����\�ɂ����Ă���B���̗������̐��]���̕��q�@�\���ǂ̂悤�Ȃ��̂��͑�ϋ����[�����A�ŋ߂̌����ł́A�����A�������̐��B�B�h���z��������e�̈�`�q�̔����̃I���^�I�t�̒��߂��Y���A�������ꂼ��ōs�Ȃ��邱�Ƃɂ���Đ��B�B�̋@�\�I���]�����\�ɂ����Ă��邱�Ƃ����炩�ƂȂ���[3]�B����A�Y�@�\���A���@�\�����ꂼ��ɂ����闼���B�B�̋@�\�ω��̎��Ԃ���ъ��v���Ƃ̊ւ��A�@�\�I���]���ߒ��ɂ�����]�̐��������ǂ̂悤�ɂȂ��Ă��邩�͋����[���B
������
���I�Y���͊��S���A�����I���͂Ƃ������Ғœ�����ʂɂ݂��錻�ۂł���B����܂ŁA��ɋ��ނŗl�X�Ȍ��ۂƂ��ĕ���Ă������A���̎d�g�݂͍ŋ߂ɂȂ�܂ŁA�قƂ�Ǖ������Ă��Ȃ������B�ŋ߂̕��q�����w�I��@��p������͂��炻�̎d�g�݂����X�ɖ��炩�ɂȂ����[2]�B���ނł́A��U�A�Y���邢�͎��Ƃ��čs������̂��t�̐��Ƃ��čs�����邱�Ƃ��\�ł���B�܂�A���ނł͈�U�A�]���Y�^���邢�͎��^�ɕ������Ă��������ɂ���ėe�Ղɋt�̐��ɐ��]�����邱�Ƃ��ł��邱�Ƃ��Ӗ�����B����́A�M���ނ̂悤�ɔ]�̐�������́A���]�����Ȃ��̂Ƃ͖��炩�ɈقȂ�B���ނł̐��I�Y���̃��J�j�Y���𖾂̂��߂̌����͋��ނ݂̂Ȃ炸�A�q�g�ł݂��鐫�����Q�̂悤�Ȏ����𖾂ɍv���ł���̂͂Ȃ����Ǝv���A���ނ̔]�̐������@�\���ǂ̂悤�ɂȂ��Ă��邩������̏œ_�̈�ł��邱�Ƃ͋^�����Ȃ��B�������A���I�Y���̕��q�@�\�̉𖾂̌����͂��̐��N�Œ������i�W�������Ă��邪�A���ʂ̊W�ō���͂��̏ڍׂɐG��邱�Ƃ͂ł��Ȃ������B�܂��ʂ̋@��ɏЉ�����B
�Q�l����
- ���ނ̐�����Ɛ��B���̐������^���]���@In�F�����̐��͂ǂ̂悤�Ɍ��܂邩�H �`���� �j�_ �y�f Vol. 49, pp. 116-123. �����o�� (2004)
- Molecular aspects of gonadal differentiation in a teleost fish, Nile tilapia. Sexual Development, 3 (2-3), 108-117. Review. (2009)
- Sex-change in the Gobiid Fish is mediated through Rapid Switching of Gonadotropin Receptors from Ovarian to Testicular Protion or Vice-versa. Endocrinology, 150 (3), 1503-1511. (2009)
�i�ȏ�j
�������̃R�����F�@-��25��-�@�u��R�J�[�e���Ɏg�p���ꂽ�L�f�n��R�܂̃��X�N�]���v�@�O�� �S��
�����ЂƂO�̃R�����F�@-��23��-�@�u�z�����A���f�q�h�̃��X�N�]���ɂ��Ă�2010�N���݂̏��v�@�J�J �h�j

