
メダカ近縁種を用いて多様化した性決定の仕組みを解明
助教 明正 大純 (Assist. Prof. Taijun Myosho)生態発生遺伝学研究室
はじめに
私たちが普段、目にする生物は、元をたどれば単細胞生物から出発し、連続的に変化してきました。その中で特定の生物が繁栄することで、非常に多様な生物相を形成しています。生物の進化は一本の樹に例えることができます。大樹の幹から枝を出し、枝分かれを繰り返す種の多様化とランダムに枝が切り落とされる種の絶滅の過程を経ます。幹から枝の先端までが進化の歴史であり、たどり着いた枝の先端が現行種を表します。この連続的な変化は、形態・構造・行動・生理的性質などの変化に密接に繋がるDNAの塩基配列の突然変異です。これらを積み重ねることによって、数百万種類に及ぶ多様な生物相を形成してきました。つまり、生物の違いはDNAの塩基配列の違いであり、その違いは生殖によって世代を重ねることによって受け継がれてきました。
多様な性決定遺伝子の同定
有性生殖、つまり雄と雌の存在は、種の維持または連続的な変化を途絶えさせないために、現在では多くの種で採用されています。その中でも多数派を占める遺伝的な性決定の仕組みは古くから脊椎動物において研究されてきました。1990年にヒトの性を決める最初のスイッチを入れる遺伝子(性決定遺伝子)であるSRYが同定されました。しかし、SRYは哺乳類で共通の性決定遺伝子であることが確認されたものの、他の生物種からSRYは発見されないことから、SRYとは異なる性決定遺伝子の存在が示唆されました。その後、2002年にメダカにおいて、脊椎動物で2番目の発見となる性決定遺伝子Dmyが同定されました。さらに、メダカ属の近縁種を調べたところ、数種の近縁種のみしかDmyを持たないこと、つまりDmy以外の分子が性決定遺伝子であることが分かりました (Matsuda et al. 2002; 2003)。Dmyを皮切りに、これまでに鳥類・両生類・魚類で10の性決定遺伝子またはその候補が同定されていますが、これらの結果は、性決定遺伝子の多様性を支持し、性決定遺伝子が頻繁に交代しうることを示唆しました。
性決定遺伝子の交代機構を解明することで、多様な性決定遺伝子がなぜ存在するかを理解することができます。性決定遺伝子の交代は2つの機構から成り立っています。1つ目は、個体レベルで起こる「性決定遺伝子の性決定機能獲得」であり、2つ目は個体群レベルで起こる「旧性決定遺伝子との交代」です。筆者は、これまでにメダカ属の近縁種であるルソンメダカの性決定遺伝子を同定することにより、上記2つの機構の一端を解明してきましたので、ここで紹介します。
ルソンメダカの性決定遺伝子GsdfY

体色に関連する5つの遺伝子の突然変異を組み合わせたメダカ。白矢印のヒメダカが♂、その右のシロメダカが♀。
メダカ属魚類は20種以上から構成され、日本の青森県を北限に東アジアに分布しています。日本に生息するメダカはミナミメダカ (Oryzias latipes)とキタノメダカ (O. sakaizumii)であり、ミナミメダカのヒメダカとシロメダカを用いて、黄色の体色を決定する遺伝子RがY染色体上にあるという性に関する研究結果(限性遺伝)が今から100年ほど前に報告されています(Aida 1921)(図1)。現在では、上記2種に加えてチュウゴクメダカ (O. sinensis)、ハイナンメダカ (O. curvinotus)の4種の性決定様式はXX♀、XY♂になるXX-XY型、性染色体は1番染色体であり、Y染色体上にあるDmyを性決定遺伝子として用いていることが判明しています (図2)。Dmyはこの4種の共通祖先で性決定機能を獲得し、旧性決定遺伝子からDmyに交代したことになります。しかし、Dmyを持つ4種の共通祖先の子孫であるルソンメダカ (O. luzonensis)はDmyを持っていません。この種では、Dmyから新規性決定遺伝子への交代が起こったと考えられます。この新規性決定遺伝子は現在までに同定された性決定遺伝子の中で最も新しく、新規性決定遺伝子の誕生の原因となった突然変異以外の突然変異の蓄積が少ないことを意味します。つまり、新規性決定遺伝子の誕生の原因となった突然変異を同定しやすいと考えられます。そこで、ルソンメダカの性決定遺伝子を同定することによって、「新規性決定遺伝子の性決定機能獲得」と「Dmyから新規性決定遺伝子への交代機構」の解明を試みました。

ルソンメダカの新規の性決定遺伝子は誕生して間もない。
性連鎖解析によって、ルソンメダカの性決定様式はXX-XY型で、性染色体はメダカの12番染色体に相当することが判明しました(Tanaka et al. 2007)。さらに、ポジショナルクローニングと発現解析により、Gsdfが性決定遺伝子の最有力候補になりました。Gsdfは性染色体上に対立遺伝子としてGsdfY(Y染色体上のGsdf)とGsdfX(X染色体上のGsdf)があります。ルソンメダカのXX個体にGsdfYを組み込んだ遺伝子導入メダカを作成したところ、導入遺伝子を持つ個体はXX♂に分化しました。一方で、GsdfXを遺伝子導入した個体ではXX♂への分化はみられませんでした。以上の結果から、GsdfYが新規に性決定機能を獲得した性決定遺伝子であることを示すことができました(Myosho et al. 2012)。
性決定遺伝子GsdfYの性決定機能獲得の機構
ルソンメダカのGsdfには、対立遺伝子としてGsdfYとGsdfXがあります。GsdfYが性決定機能獲得に至った原因配列を探るために両者を比較しました。遺伝子がタンパク質として働くアミノ酸配列と遺伝子が機能する場所であるmRNAの発現する細胞に違いはなく、GsdfYはGsdfXより早く発現し始めることが唯一の違いでした。そして、GsdfYの上流のエンハンサー配列の数塩基の変化によって、この早い発現が誘導されることを支持する結果が得られました。
Gsdfは細胞の増殖や分化に関わるTGF-βスーパーファミリーの一員であり、ニジマスで初めて同定された魚類特有の遺伝子です(Sawatari et al. 2007)。Dmyを性決定遺伝子に持つ4種のメダカでは性分化時期に雄特異的な高発現を示します。性決定遺伝子が最初にスイッチを入れ、最終的に精巣が形成されるまでに順番に遺伝子が機能していくという性決定の遺伝子カスケードで考えると、GsdfはDmyの制御下にあり、そしてDmyより後で働く性分化遺伝子であるといえます。以上の結果から、ルソンメダカのGsdfYは、性決定カスケードにおいてDmyの下流で機能していたGsdfがエンハンサー配列の変化によって性決定機能を獲得したことが強く示唆されました(図3)
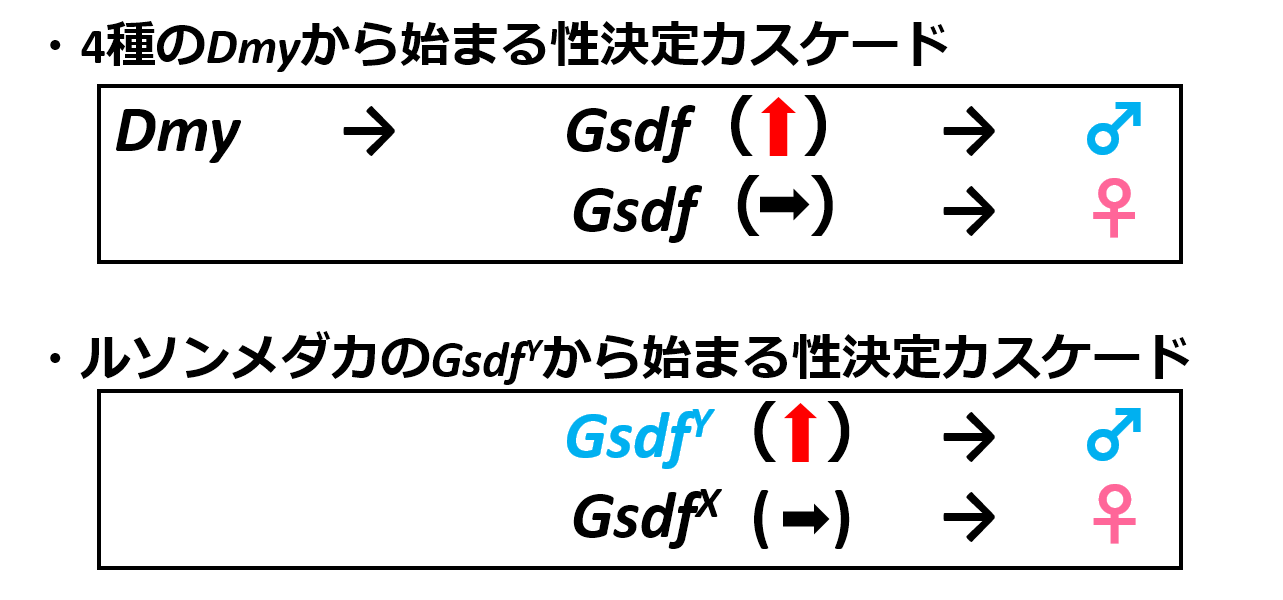
ルソンメダカの性決定カスケードではGsdfYがDmyに依存しない高発現を獲得することで、性決定機能を獲得した。
DmyからGsdfYへの性決定遺伝子交代のシナリオ
「DmyからGsdfYへの交代機構」を解明するために、ミナミメダカの雌(XX)にルソンメダカのGsdfYを導入したところ、導入遺伝子を持つXX♂個体が得られました。この結果は、DmyがなくてもGsdfの発現を誘導することによって正常な雄分化過程をたどること、Gsdfより下流の性決定カスケードはミナミメダカとルソンメダカで共通であることを示唆しました。さらにこの結果は、性決定遺伝子の交代には、性決定遺伝子の下流の性決定カスケードの変更を必要としないことを意味しています。
全ての結果をまとめると、以下の性決定遺伝子交代のシナリオが考えられます(図4)。5種の共通祖先では、Gsdfの発現を上げるDmyが唯一の性決定遺伝子として雄決定を担っていました。次に、ルソンメダカの祖先において、GsdfがDmyに依存しない高発現能による性決定機能を獲得したことで、第2の性決定遺伝子であるGsdfYが個体群中に誕生しました。次に、遺伝的浮動やボトルネック効果などが原因で個体群中のGsdfYの遺伝子頻度が高まり、 2つの性決定遺伝子DmyとGsdfYが共存する状態になりました。最後に、Dmyが個体群中から失われることによって新規性決定遺伝子GsdfYへの交代が完了した今のルソンメダカになったと考えられます。
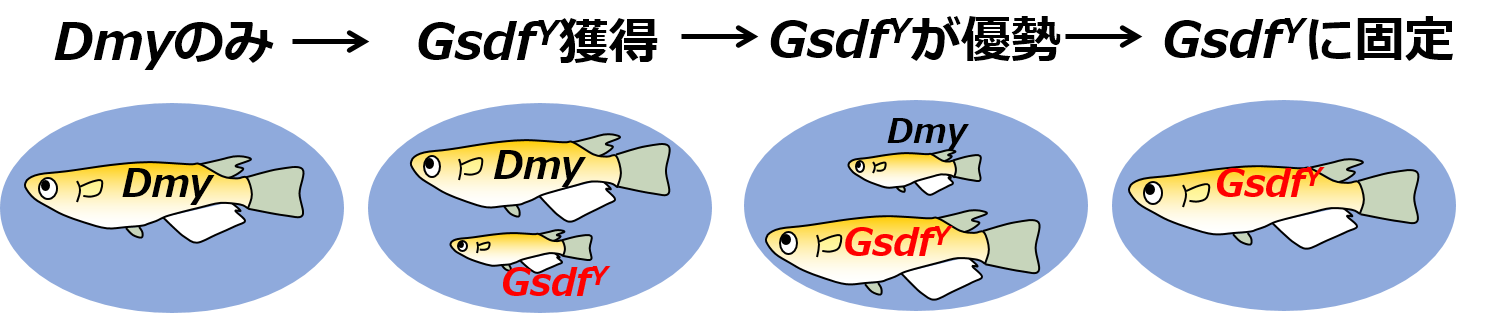
個体群中の♂のみを示し、メダカの大きさが個体数を表す。
おわりに
ルソンメダカの性決定遺伝子の同定によって、旧性決定遺伝子Dmyから新規性決定遺伝子GsdfYへの交代機構を推定することができました。現在まで同定されている、そしてこれから見つかる多様な性決定遺伝子も同様のプロセスを経ることによって誕生したと考えらえます。今回の結果は性決定遺伝子の交代機構の推定に留まりました。今後は、日本に生息するミナミメダカまたはキタノメダカの野生個体群の解析によって、性決定遺伝子が交代する瞬間である性決定遺伝子が共存する個体群を探索し、多様な性決定遺伝子を持つ現行種への理解を深めていきたいと考えています。
参考文献
- . Nature 417, 559-563, 2002.
- . Zoolog. Sci. 20, 159-161, 2003.
- Genetics 6, 554-673, 1921.
- . Genetics 177, 2075-2081, 2007.
- . Genetics 191, 163-170, 2012.
- . Dev. Biol. 301, 266-275, 2007.
(以上)


{kind=link}
{kind=link}